Section 1. Results of Prior Support
The research team for the initial 6-year phase of the McMurdo Dry Valleys LTER project (MCM-I)
consisted of R. A. Wharton, Jr. (lead PI/limnologist), A. G. Fountain (glaciologist), D. W.
Freckman (now D. H. Wall) (soil ecologist), W. B. Lyons (geochemist), D. McKnight (stream
ecologist/hydrologist), D. L. Moorhead (ecological modeler), J. C. Priscu (limnologist) and C.
M. Tate (stream ecologist). During MCM-I, we produced 68 papers in refereed journals, 32 book
chapters, and 8 unrefereed journal articles, 7 dissertations and theses, over 95 abstracts from
national and international meetings, and 37 papers in the NSF-Office of Polar Programs publication,
Antarctic Journal of the United States. We supported 28 graduate students and 7 post doctoral
fellows. A total of over 60 collaborators have been involved in MCM-I research. For more details,
see the McMurdo LTER publications web page.
Results of the initial 6-year phase of the McMurdo Dry Valleys LTER project recently have been
compiled and published in two synthesis volumes
(1) Ecosystem Processes in Antarctic Ice-free
Landscapes (Lyons et al. 1997) and (2)Ecosystem Dynamics in a Polar Desert: The McMurdo Dry Valleys, Antarctica (Priscu 1998,
American Geophysical Unions Antarctic Research Series). These two books represent our first
attempts at integration and present an overview and synthesis of ecosystem processes within this
extreme environment, including interactions between physical, chemical, and biological components.
In both books, the dry valley regions of Antarctica are presented within an ecosystem
context representing a significant departure from earlier subject-specific compilations of Antarctic
dry valley studies (Pickard 1986; Green and Friedmann 1993; Bormann and Fritzsche 1995). A CD-ROM
also accompanies the Priscu volume and provides detailed geospatial data to support the text.
Prior to MCM-I, research in the MCM was sporadic, discipline-specific and lacked an integrated ecological
basis. Among the most important contributions of MCM-I was the integration of these earlier site-specific
studies on streams, lakes, glaciers, and soils, into an ecosystem perspective of Taylor Valley (the principle
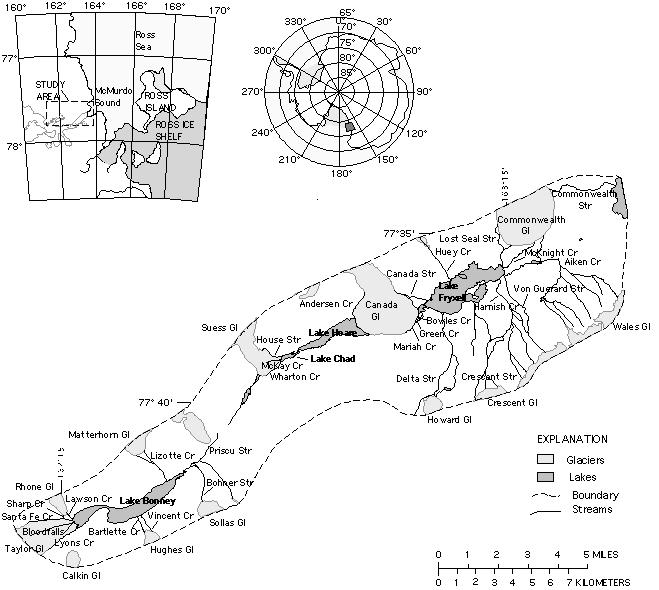
site of MCM-I investigations; Fig.
1.1). This framework has proven critical to evaluating both interactions among various elements of the
MCM ecosystem and their responses to current and projected environmental change. Our work in addressing the
LTER core areas of research and developing a modeling framework to simulate the functioning of the MCM
ecosystem, provided a means of predicting ecosystem behavior. Below we summarize the main results of our
research on each of the major landscape units (lakes, streams, soils, glaciers) including our progress in
modeling and synthesis.
Fig. 1.1 TAYLOR VALLEY, southern Victoria Land, Antarctica, primary field location
of MCM-I and MCM-II.
Meteorology:
Climate drives and shapes all ecological systems, so all LTER sites include some degree of
meteorological monitoring. However, MCM requires a particularly large meteorological program
because no larger network of meteorological stations exists in the region, meteorological conditions
show a high degree of spatio-temporal heterogeneity, and biological activities are particularly
sensitive to microclimatic conditions. Our LTER Automatic Weather Network (LAWN) consists of eleven
stations (Fig. 1.2) collecting all the standard meteorological variables as well as specific
measures particularly important to the MCM (Doran et al. 1995).
Significant differences in the climate regime in MCM occur over very short distances. For instance,
the three main Taylor Valley lake basins lie within 20 km and 47 m elevation, but Lake Bonney
experiences a continental climate driven by katabatic winds and Lake Fryxell experiences a maritime
climate (
Table 1.1).
The climate of Lake Hoare is transitional between the two, possibly because
the Nussbaum Riegel (700 m high hill in the center of Taylor Valley) blocks moisture-bearing clouds
from the ocean from reaching the Lake Bonney basin (Fountain et al. 1998; Lyons et al. in review
b). This landscape feature controls the pattern of precipitation, humidity, and winds in Taylor
Valley, which partly defines ecological conditions.
Table 1.1. Average meteorological conditions within three
lake basins of Taylor Valley during summer (1994-96).
|
Basin
|
Temperature (�C)
|
Relative Humidity (%)
|
Wind Speed (m/s)
|
Solar Radiation (mmol/m�/s)
|
|
Fryxell
|
-7.7
|
59.3
|
3.4
|
76.0
|
|
Hoare
|
-7.4
|
59.7
|
2.8
|
57.0
|
|
Bonney
|
-6.7
|
53.3
|
4.6
|
45.6
|
The meteorological data have been used in most aspects of MCM-I research, such as predicting glacial
melt (Dana et al. 1998) and streamflow (Conovitz et al. 1998; Lewis et al. in press, a; Fountain
et al. 1998), and as drivers for soil ecosystem dynamics (Treonis et al. 1997) and ecological
modeling (Moorhead and Priscu 1998). Indeed, micro-meteorological conditions have important
implications for the distribution and productivity of the biological communities in terrestrial and
aquatic environments. For example, low humidity and snow results in more arid and saline soils and
limited soil biota in the Lake Bonney basin compared to the Lake Fryxell basin. Less snow cover
within the Bonney basin also reduces albedo, which increases the melt water flux from glaciers and
reduces interannual variability in streamflow (Fountain et al. 1998). Spatio-temporal patterns of
solar radiation within Taylor Valley are strongly influenced by topography (Dana et al. 1998), and
are modified by lake ice thickness and snow cover. The pattern in solar radiation influences primary
production in the Taylor Valley lakes (Lizotte and Priscu 1998).
Glaciers:
Our long-term program of glacier measurements is unique among LTER sites, and a critical component of
the MCM project because melting glaciers provide the bulk of water to the streams and lakes in the
MCM (Chinn 1993). For any given elevation, the annual mass balance decreases with distance away from
the ocean in a manner consistent with the meteorological gradient (Fountain et al. in review).
Changes in glacier mass define the magnitude of ice lost to evaporation and melt water to the streams
and lakes. Results show that the glaciers have been increasing in mass since 1993, the start of
MCM-I, which coincides with a period of cooler than normal summers and more than average snowfall.
The larger glaciers seem to be advancing, but this advance is primarily a legacy of past climate,
roughly 1000 years BP (Fountain et al. 1998).
To predict melt water flow from the glaciers, field measurements of the energy balance are collected
from the glacier surface; 40-90% of the mass from the ablation zone (lower elevations) is lost to
evaporation/sublimation but the remainder is lost as melt water (Lewis et al. in press, a).
Previous studies concluded that air temperature was the dominant factor in controlling melt (Wharton
et al. 1993). However, our work shows that snow cover is equally important and controls the
variability in streamflow from different glaciers (Lyons in review b). Snow cover reflects solar
energy that otherwise would be absorbed by ice. When snow cover exists during the early and late
portions of summer, ice cliffs that form on the lower margins of the larger glaciers become important
water sources that are critical to maintaining streamflow (Conovitz et al. 1998; Lewis et al. in
press b).
Streams:
The MCM streams flow during the summer and some contain abundant algal mats, persisting in a freeze
dried state in winter. Compared to other stream ecosystems in the LTER network, MCM streams
represent several extremes because of their lack of allochthonous organic inputs, high standing
algal biomass, low primary productivity and low grazing losses (Webster and Meyers 1997; McKnight
and Tate 1997). We have documented the range of productivity by mapping the distribution of algal
mats at sites in 11 streams in Taylor Valley (McKnight et al. 1998; Alger et al. 1997). High
algal abundance occurs in moderate gradient streams with a stable stone pavement in the streambed.
In streams with sparse mats, parafluvial seeps draining the hyporheic zone are important habitats
(McKnight et al. 1998).
We established a gauging network to monitor flow of major streams in Taylor Valley, and have shown
that flow patterns are controlled by glacier melt and stream geomorphology, including water storage
in the hyporheic zone (saturated area beneath and adjacent to the stream) (Von Guerard et al. 1995;
Conovitz et al. 1998). Tracer experiments indicate rapid hyporheic exchange in MCM streams (Runkel
et al., in press). Major cation and silicate data indicate chemical weathering generates solutes in
the hyporheic zone and that weathering rates are high (Lyons et al. 1997b, 1998).
Previous work focused on the biogeochemical evolution of the lakes, but our data demonstrate that
instream processes also influence lake chemistry (Moorhead et al. 1998). Monitoring of stream
chemistry showed that nutrient concentrations are higher in streams without mats than in streams
with mats (Table 1.2).
We conducted a nutrient injection in a stream with abundant algal mats with
injectate concentrations corresponding to drainage of nutrient-rich hyporheic water in late summer
(55 M and 18 M for NO3(-) and PO4(3-), respectively) (McKnight et al. in review a). By 497 m below the
injection, nutrients remained below detection (<1-2 M) during the arrival of the Cl tracer,
illustrating rapid nutrient uptake. At two intermediate sites, production of NO2(-) and NH4()+
indicated occurrence of dissimilatory nitrate reduction in the hyporheic zone. We used a solute
transport model with nutrient uptake represented as a first order process to determine reach scale
parameters for N and P uptake. The best match to the experimental data was a model in which P
uptake occurred in the main channel and N uptake occurred in the main channel and in the hyporheic
zone, representing the loss due to dissimilatory nitrate reduction (7-16% of total N uptake).
Table 1.2. Visual Algal abundances for Taylor Valley Streams and
related NO3 and PO4 values.
["Low" denotes <50% surface cover; "high" denotes >50% surface cover;
ND denotes "no data available"]
|
Stream and Basin
|
Total Stream Length (km)
|
Visual Algal Abundance
|
(Sites nearest
NO3 (uM)
|
the outlet)
PO4 (uM)
|
|
Fryxell Basin
|
|
Huey Creek
|
2.1
|
low
|
4.62
|
0.40
|
|
Canada Stream
|
1.5
|
high
|
0.74
|
0.25
|
|
Bowles Creek
|
0.9
|
high
|
0.71
|
0.21
|
|
Green Creek
|
1.2
|
high
|
0.79
|
0.15
|
|
Delta Stream
|
11.2
|
high
|
0.81
|
0.09
|
|
Von Guerard Stream
|
4.9
|
high
|
0.89
|
0.42
|
|
Hoare Basin
|
|
Andersen Creek
|
1.4
|
low
|
4.17
|
0.23
|
|
House Creek
|
2.0
|
low
|
3.91
|
0.65
|
|
Wharton Creek
|
1.0
|
low
|
ND
|
ND
|
|
Bonney Basin
|
|
Priscu Stream
|
3.8
|
low
|
5.98
|
0.36
|
|
Lawson Creek
|
0.3
|
low
|
12.17
|
0.21
|
|
Bohner Stream
|
1.9
|
low
|
9.28
|
0.61
|
Monitoring of stream chemistry has shown that the streams have high HCO3(-):DOC molar ratios (~4.5)
compared to ratios of 1.0-1.6 for Arctic rivers (Gordeev et al. 1966; Cauwet and Sidrov 1996) and
average ratios of 3.5 for rain forest streams (McDowell and Asbury 1994; LUQ). The higher HCO3(-):DOC
ratios in MCM streams reflect the lack of terrestrial organic carbon input, the interannual
preservation of freeze-dried algal mats and high weathering rates producing HCO3(-) (McKnight et al.
1991; Lyons et al. 1998). Measurements of stream suspended load (SSL) have shown that total SSL
from the Fryxell and Hoare basins are much lower than those from the Bonney basin
(Table 1.3).
Stream DOC concentrations are more variable than particulate organic matter (POM) concentrations,
and streams in Fryxell basin have higher DOC:POC ratios than those in Bonney basin. The Fryxell
ratios are similar to those of tundra (2.6) and tropical (1.1) fluvial systems, while those of
Bonney are closer to warm desert rivers/streams (0.4) (Ludwig and Probst 1996). Such differences
in the partitioning of organic carbon reflect fundamental differences in carbon transport between
basins.
Table 1.3. Sediment load, POC
and DOC loading (mg/L) from Taylor Valley streams.
|
Stream
|
Suspended Load
|
POC
|
DOC
|
DOC/POC
|
|
Fryxell Basin
|
|
Green
|
1.4
|
0.6
|
1.25
|
2.1
|
|
Aiken
|
8.4
|
1.2
|
2.08
|
1.7
|
|
McKnight
|
4.2
|
1.0
|
1.50
|
|
|
Bowles
|
4.6
|
1.2
|
1.35
|
1.12
|
|
Mariah
|
17.8
|
0.7
|
0.53
|
|
|
Hoare Basin
|
|
Anderson
|
13.0
|
0.6
|
0.47
|
0.78
|
|
Bonney Basin
|
|
Lawson
|
52
|
1.3
|
0.47
|
0.47
|
|
Lyons
|
293
|
1.45
|
1.81
|
0.33
|
|
Santa Fe
|
362 +/- 30
|
2.7
|
0.68
|
0.48
|
|
Priscu
|
69
|
0.7
|
|
|
To understand the long-term persistence of stream algal mats, we studied a relict stream channel for
which sustained flow was last recorded in the summer of 1969 (McKnight et al. in review b). We
routed meltwater to the channel and found that relict algal mats began growing within a few days at
rates exceeding those of mats in streams with regular summer flow, because of greater solute and
nutrient concentrations. This long term experiment of MCM-I has shown that cryogenic preservation
of algal mats in inactive channels allows for rapid response to climatic and geomorphological
shifts.
Lakes:
Although many lakes in the LTER sites develop winter ice-cover, lakes in the MCM are unique among
LTER sites because they are covered by 3-6 m of perennial ice. This ice reduces light penetration
and circulation, alters sediment pathways, severely reduces mixing, and restricts gas exchange with
the atmosphere. Planktonic food webs also are unique in that they are limited to algae, bacteria,
protozoans and rotifers (Laybourn-Parry et al. 1997; James et al. 1998). Our studies have
provided fundamental information on the physical, chemical, and biological attributes of the three
main lakes (Bonney, Hoare, and Fryxell) in Taylor Valley.
Changes in lake volumes represent regional climate change in the recent past, as lake levels
generally have been rising from the 1960s to 1992, and Lake Bonney has been rising since 1905 (Chinn
1993). Measurements from 1993-98 indicate that lake levels are no longer rising significantly.
These observations are consistent with measurements of streamflow and glacier mass balance and
result from cooler temperatures and heavier snows during summers. The lack of physical mixing leads
to stable physical and chemical stratification. The geochemistries of the surface waters and
monimolimnia of the lakes are distinct
(Table 1.4), implying
differences in glacier source, stream length, stream channel geology, and different developmental
histories, respectively (Lyons and Welch 1997; Lyons et al. 1998; Lyons et al. in review a).
Table 1.4. Chemical characteristics
of Taylor Valley lakes, September 1995.
|
Lake
|
Depth (m)
|
Li (M)
|
Na (mM)
|
K (mM)
|
Mg (mM)
|
Ca (mM)
|
Cl (mM)
|
Br (mM)
|
SO4 (mM)
|
DIC (mmol)
|
|
Hoare
|
5
|
0.7
|
2.47
|
0.27
|
0.22
|
0.75
|
2.2
|
0.0022
|
0.42
|
1.72
|
|
Hoare
|
30
|
2.3
|
7.81
|
0.77
|
1.4
|
2.14
|
6.3
|
0.0062
|
0.92
|
8.18
|
|
Fryxell
|
5
|
1.6
|
7.9
|
0.54
|
0.99
|
1.35
|
7.59
|
0.009
|
0.45
|
4.68
|
|
Fryxell
|
18
|
16
|
117
|
4.79
|
13
|
3.76
|
99
|
0.141
|
1.58
|
48
|
|
W Bonney
|
5
|
3.1
|
10
|
0.36
|
1.47
|
1.79
|
12
|
0.022
|
1.54
|
0.91
|
|
W Bonney
|
38
|
656
|
1789
|
43
|
402
|
62
|
2299
|
5.06
|
47
|
78
|
|
E Bonney
|
5
|
3.6
|
12
|
0.381
|
1.78
|
2.05
|
14
|
0.028
|
1.73
|
1.41
|
|
E Bonney
|
39
|
1169
|
2701
|
67
|
1247
|
32
|
5100
|
19
|
36
|
3.66
|
Our data indicate that respiration exceeds primary production in the water column of MCM lakes
(Lyons et al. in review b), despite extremely low inputs of allochthonous organic carbon (McKnight
et al. 1993; Aiken et al. 1996). Thus, MCM lakes are net heterotrophic systems, much like other
unproductive aquatic systems (del Georgio et al. 1997). The source of excess carbon is a large
pool of organic carbon that appears to be a legacy of a previous stage in lake development (Aiken et
al. 1996), as indicated by 14C dating of the POC pool (Doran 1996). The surface pool of POC in
Lake Bonney is 10,000-13,500 yrs old (Doran 1996), but only 20% of it is refractory humic materials
(McKnight et al. 1991). Therefore, the carbon dynamics of the MCM lakes are different than those
of other lakes in the LTER network (NTL, HBR, NWT and ARC), and may be comparable to some lakes in
Siberia where such legacy carbon also is being utilized as an energy source by modern communities
(Zimov et al. 1997).
Phytoplankton communities in MCM lakes exist in an environment characterized by low light intensity.
Light intensity and spectral composition vary with depth and phytoplankton occur in relatively
distinct, stratified layers (Spaulding et al. 1994; Lizotte and Priscu 1998). A considerable
degree of shade adaptation exists among phytoplankton (Neale and Priscu 1998). Vertical nutrient
profiles suggest that deep maxima of primary production are driven by diffusion of nutrients from
even deeper water, where nutrient pools have accumulated as a legacy of past stages in lake
development (Priscu 1995). Although these systems lack many of the grazers found in aquatic
communities, grazing by microzooplankton and protozooplankton may affect phytoplankton communities,
and recent investigations are beginning to identify and quantify the taxa comprising planktonic food
webs (Laybourn-Parry et al. 1997; James et al. 1998). An interesting discovery of MCM-I is that
mixotrophic phytoplankton species have been identified and phagotrophy observed. Mixotrophy
represents a survival strategy in that phytoplankton may utilize heterotrophic metabolism during
prolonged periods of low light conditions (Moorhead and Priscu 1998). A study of the over-winter
dynamics of phytoplankton in Lake Fryxell (Fig. 1.3) showed that mixotrophic species increased in
abundance (McKnight et al. 1998). Large concentrations of virus-like particles have been found in
Lakes Hoare and Fryxell (Kepner et al. 1997), suggesting a possible role of viruses as regulators
of microbial community dynamics (Fig. 1.4).
MCM lakes also support abundant growths of benthic microbial mats. These mats are dominated by
cyanobacteria and, though acclimated to the continuously low light environment, are always light
limited. Light limitation has been used to explain the decrease in mat biomass with depth in Lake
Hoare (Wharton et al. unpublished; Moorhead et al. 1997b). Lacustrine organic matter,
representing a legacy of past productivity, is found in soils, perched deltas on the valley walls,
and former lacustrine sand mounds on the valley floors. In modern times, organic matter does
accumulate in the surface sediments of Lakes Fryxell and Hoare, mostly from benthic microbial mat
production (Lawrence and Hendy 1985; Anderson et al. 1993; Doran et al. in review).
Soils:
The MCM serve as a model system for elucidating the ecological roles of soil biota. Globally, there
are no other soil systems where nematodes represent the top of the food chain and where food webs
have such simple structure. The food webs in the soils are limited largely to algae, yeasts,
bacteria, protozoans and nematodes (Freckman and Virginia 1997a, 1998). The majority of soils
sampled across the valleys (65%) support up to three soil invertebrate taxa (tardigrades, rotifers,
nematodes), although other sites show a complete lack of these invertebrates. Protozoa also
contribute to the diversity of soil communities and may compete with the endemic microbial-feeding
nematode, Scottnema lindsayae (Bamforth et al. 1996). Despite their trophic dominance, the species
diversity of nematodes is very low (n = 3), representing only 2 functional levels (predator and
microbivore) compared to 5 functional groups more common in other systems (e.g. JRN, SEV, CSGS,
BNZ).
We investigated the distribution and functional significance of these low biomass and low diversity
soil communities throughout the MCM. This information is being compared to our knowledge of soil
biotic function in other arid systems, including those of the LTER network (JRN, SEV, CSGS). For
example, detailed examinations of the survival and community ecology of soil nematodes in the MCM
show that, in contrast to hot deserts where plant-related factors limit soil biotic communities,
soil chemical factors may be most important in defining location and structure of communities
(Freckman and Virginia 1997). There is no single soil property that defines a suitable or
unsuitable habitat, but where soil biodiversity is reduced, soil physical and chemical heterogeneity
is high (Freckman and Virginia 1997). We have found that ribosomal and mitochondrial DNA of
Scottnema lindsayae varies across small (<1 m2) and large (60 km2) spatial scales, suggesting that
the species may be evolving (to date, 11 distinct maternal lineages have been detected in the MCM,
Courtright et al. in press). This evolution may be a response to the high spatial heterogeneity of
the soil environment (Ho et al. 1995).
The simple food chains found in MCM soils appear to be strongly influenced by human disturbance. A
long-term manipulation experiment shows that soil warming and increased moisture and carbon
availability have major effects on the nematode community, decreasing the abundance of the
omnivore-predator species, increasing the abundance of a microbivorous species (Freckman and
Virginia 1997), and altering soil respiration (CO2 efflux). We presently are examining how
community composition influences decomposition (Treonis et al. 1997; Burkins et al. 1997).
We have developed an initial soil carbon budget for the MCM based upon systematic regional sampling
of soil profiles (Brown et al. 1996; Burkins et al. 1997). The natural abundances of 13C and 15N
in soil organic matter indicate that relative contributions of marine, soil derived, and lacustrine
(recent and legacy) sources to soil pools is a function of location in the MCM landscape (elevation;
distance from lakes, streams, or paleolakes; distance from marine sources). Relationships between
soil community structure, biological activity and quantity and source of soil organic matter suggest
that the soil carbon cycle in the MCM represents an extreme end-member of global soil ecosystems,
with an extraordinarily slow rate of C-cycling, perhaps comparable to Arctic peat.
Ecological Modeling:
Our initial view of the MCM ecosystems was that biological communities were controlled primarily by
temperature and hydrologic regimes, as they determine the availability of liquid water. However,
energy availability likewise controls the location, structure and activity of MCM communities
(Moorhead and Priscu 1998). Paradoxically, microsites with exposure to radiant energy also may be
subject to rapid desiccation and potentially rapid freeze-thaw cycles that limit biological
activity. A generalized ecosystem model has been developed to simulate energy and nutrient dynamics
in MCM communities (Fig. 1.5). Organic matter may accumulate through on-site photosynthesis or
allochthonous inputs, and be lost through on-site respiration, release of dissolved organic carbon
or mechanical erosion by wind and water. However, ecosystems within the MCM differ with respect to
the importance of various environmental controls. Because the activity of biota are restricted to
those times and places with a favorable juxtapositio n of energy and moisture regimes, measurements
of microclimate and energy availability needed to drive simulation models of primary and secondary
productivity must be available at high resolution in both space and time. For these reasons,
modeling activities of MCM I have focused on aquatic environments, for which patterns of moisture
availability are known with some certainty.
To date, we have developed models to simulate primary production of benthic microbial mats found in
stream and lake beds (Moorhead et al. 1997a, 1997b, 1998), as well as plankton communities in MCM
lakes (Davis 1988). Photosynthesis of these communities saturate at low-light intensities, and
Moorhead et al. (1997b) explored the ramifications of using different models of photosynthesis
(rectangular hyperbolic, hyperbolic tangent and linear response models). We found that because
stream mats usually are light-saturated, only maximum rates of photosynthesis affected simulation
results. However, lake mats receive such low levels of radiant energy (Howard-Williams et al.
1998) that differences in low-light responses of the various models had substantial impacts on model
behavior. These results demonstrated the critical need for monitoring light regimes within the
water columns of MCM lakes at higher spatial and temporal resolution.
Results of our modeling studies suggest a positive, net annual primary production of microbial mats
in streams and shallow depths in lakes, in part, because decomposition is slow and other
hetrotrophic activity is virtually absent. For these reasons, organic carbon accumulates,
consistent with the notion that ancient aquatic ecosystems in the MCM produced much of the organic
carbon reservoir present in modern soils (Burkins et al 1997). This modeling approach also has been
used to simulate immobilization of nitrogen by microbial mats in streams (Moorhead et al. 1998),
and has shown that N immobilization approximates that required to balance net C fixation. This
model now is being used to evaluate N and P limitations to the location and productivities of
plankton communities in MCM lakes (Davis 1998).
Legacies and Linkages:
Doran et al. (1994) and Lyons et al. (1997a) have summarized the importance of past climate
conditions on MCM and the ecological legacies that these climatic changes have created, and Priscu
(1995) has demonstrated the importance of these legacies on the current lacustrine ecosystem. A
model of how past and present variations in climate have controlled the chemical and biological
evolution of the MCM lakes has been developed (Lyons et al. in review, b). The degree to which the
individual lakes interact with their surrounding environment (i.e. landscape position) is a key to
the understanding of their past and present development. Present microclimatic variation and its
manifestation over the past 6000 years have led to the differences observed in modern lakes.
Heterogeneity characterizes organic carbon and nutrient distributions within modern landscapes as a
result of past climate regimes. The PIs are also committed to five articles in Bioscience (due June
1998) on MCM-LTER with legacy as the central theme.
PUBLICATIONS FROM LTER MCM-I
Books
- Lyons W. B., Howard-Williams C., and Hawes I. (eds.) (1997) Ecosystem Processes in Antarctic
Ice-Free Landscapes, pp. 281. Balkema.
- Priscu J. C. (ed.) (1998) Ecosystem Dynamics in a Polar Desert: The McMurdo Dry Valleys,
Antarctica, pp. 369. American Geophysical Union.
Refereed Journal Articles
- Aiken G., McKnight D., Harnish R., and Wershaw R. (1996) Geochemistry of aquatic humic
substances in the Lake Fryxell Basin, Antarctica. Biogeochemistry 34, 157-188.
- Barker K. R., Hussey R. S., Krusberg L. R., Bird G. W., Dunn R. A., Ferris H., Ferris V.
R., Freckman D. W., Gabriel C. J., Grewel P. S., MacGuidwin A. E., Riddle D. L., Roberts P.
A., and Schmitt D. P. (1994) Plant and soil nematodes: societal impact and focus for the future.
Journal of Nematology 26(2), 127-137.
- Bishop J. L., Koeberl C., Kralik C., Froschl H., Englert P. A. J., Andersen D. W., Pieters
C. M., and Wharton Jr. R. A. (1996) Reflectance spectroscopy and geochemical analyses of Lake
Hoare sediments, Antarctica: Implications for remote sensing of the Earth and Mars. Geochimica et
Cosmochimica Acta 60(5), 765-785.
- Brussaard L., Behan-Pelletier V. M., Bignell D. E., Brown V. K., Wim A. M. D., Folgarait P.
J., Fragoso C., Freckman D. W., Gupta V. V. S. R., Hattori T., Hawksworth D., Klopatek C.,
Lavelle P., Malloch D., Rusek J., Soderstrom B., Tiedje J. M., and Virginia R. A. (1997)
Biodiversity and ecosystem functioning in soil. Ambio 26(8), 563-570.
- Doran P. T., Wharton Jr. R. A., and Lyons W. B. (1994) Paleolimnology of the McMurdo Dry
Valleys, Antarctica. Journal of Paleolimnology 10, 85-114.
- Dyer M. I., Coleman D. C., Freckman D. W., and McNaughton S. J. (1993) Measuring
heterotroph-induced source-sink relationships in Panicum coloratum with 11C technology. Ecological
Applications 3(4), 654-665.
- Freckman D. W., Blackburn T. H., Brussaard L., Hutchings P., Palmer M. A., and Snelgrove P.
V. R. (1997) Linking biodiversity and ecosystem functioning of soils and sediments. Ambio 26(8),
556-562.
- Freckman D. W. and Virginia R. A. (1993) Extraction of nematodes from Dry Valley Antarctic
soils. Polar Biology 13, 483-487.
- Freckman D. W. and Virginia R. A. (1997) Low-diversity Antarctic soil nematode communities:
distribution and response to disturbance. Ecology 78(2), 363-369.
- Goldman C. R., Elser J. J., Richards R. C., Reuter J. E., Priscu J. C., and Levin A. L.
(1996) Thermal stratification, nutrient dynamics, and phytoplankton productivity during the onset of
spring phytoplankton growth in Lake Baikal, Russia. Hydrobiologia 20, 1-16.
- Klopatek C. C., O'Neill E. G., Freckman D. W., Bledsoe S. D., Coleman D. C., Crossley Jr.
D. A., Ingham E., Parkinson D., and Klopatek J. M. (1993) The sustainable biosphere initiative:
a commentary from the U.S. Soil Ecology Society. Bull. Ecol. Soc. Amer. 73, 223-228.
- Laybourn-Parry J., James M., McKnight D., Priscu J., Spaulding S., and Shiel R. (1997) The
microbial plankton of Lake Fryxell, southern Victoria Land, Antarctica during the summers of 1992
and 1994. Polar Biology 17, 54-61.
- Lizotte M. P. and Priscu J. C. (1994) Natural fluorescence and quantum yields in vertically
stationary phytoplankton from perenially ice-covered lakes. Limnology and Oceanography 39(6),
1399-1410.
- Lizotte M. P., Sharp T. R., and Priscu J. C. (1996) Phytoplankton dynamics in the stratified
water column of Lake Bonney, Antarctica. I. Biomass and productivity during the winter-spring
transition. Polar Biology 16, 155-162.
- Lyons W. B. and Welch K. A. (1997) Lithium in the waters of a polar desert. Geochimica et
Cosmochimica Acta 61(20), 4309-4319.
- Marion G. M., Henry G. H. R., Freckman D. W., Jones G., Jones M. H., Molau U., Molgaard P.,
Parsons A. N., Svoboda J., and Virginia R. A. (1997) Open-top designs for manipulating field
temperature in high-latitude ecosystems. Global Change Biology 2(Suppl. 1), 20-32.
- McKay C. P., Clow G. D., Andersen D. T., and Wharton Jr. R. A. (1994) Light transmission
and reflection in perennially ice-covered Lake Hoare, Antarctica. Journal of Geophysical Research
99, 20427-20444.
- McKnight D. M. and Andrews E. D. (1993) Hydrologic and geochemical processes at the
stream-lake interface in a permanently ice-covered lake in the McMurdo Dry Valleys, Antarctica.
Verhandlungen Internationaler Vereinigung Limnologie 25(957-959).
- McKnight D. M., Andrews E. D., Spaulding S. A., and Aiken G. R. (1994) Aquatic fulvic acids
in algal-rich antarctic ponds. Limnology and Oceanography 39(8), 1972-1979.
- McKnight D. M. and Tate C. M. (1997) Canada Stream: a glacial meltwater stream in Taylor
Valley, South Victoria Land, Antarctica. Journal of the North American Benthological Society 16(1),
14-17.
- Miller L. S. and Aiken G. R. (1995) Isotopic analysis of ice dynamics in Lake Fryxell,
Antarctica. Limnology and Oceanography 41, 966-976.
- Moore J. C., DeRuiter P. C., Hunt H. W., Coleman D. C., and Freckman D. W. (1996)
Microcosms and soil ecology: critical linkages between field studies and modelling food webs.
Ecology 77(3), 694-705.
- Moorhead D. L., Wolf C. F., and Wharton Jr. R. A. (1997) Impact of light regimes on
productivity patterns of benthic microbial mats in an Antarctic lake: A modelling study. Limnology
and Oceanography 42, 83-91.
- Neale P. J. and Pricsu J. C. (1995) The photosynthetic apparatus of phytoplankton from a
perenially ice-covered Antarctic lake: acclimation to an extreme shade environment. Plant Cell
Physiology 36(2), 253-263.
- Overhoff A., Freckman D. W., and Virginia R. A. (1993) Life cycle of the microbivorous
Antarctic Dry Valley nematode Scottnema lindsayae (Timm 1971). Polar Biology 13, 151-156.
- Powers L. E., Freckman D. W., and Virginia R. A. (1994) Survival of Scottnema lindsayae
under extreme osmotic conditions. Journal of Nematology 26, 116.
- Powers L. E., Freckman D. W., and Virginia R. A. (1995) Spatial distribution of nematodes in
polar desert soils of Antarctica. Polar Biology 15, 325-333.
- Priscu J. C. (1995) Phytoplankton nutrient deficiency in lakes of the McMurdo Dry Valleys,
Antarctica. Freshwater Biology 34, 215-227.
- Priscu J. C. (1997) The biogeochemistry of nitrous oxide in permanently ice-covered lakes of the
McMurdo Dry Valleys, Antarctica. Global Change Biology 3, 301-315.
- Priscu J. C., Downes M. T., and McKay C. P. (1996) Extreme supersaturation of nitrous oxide in a
poorly ventilated Antarctic lake. Limnology and Oceanography 41(7), 1544-1551.
- Robertson G. P. and Freckman D. W. (1995) The spatial distribution of nematode trophic groups
across a cultivated ecosystem. Ecology 76(5), 1425-1432.
- Spaulding S. A., McKnight D. M., Smith R. L., and Dufford R. (1994) Phytoplankton population
dynamics in perennially ice-covered Lake Fryxell, Antarctica. Journal of Plankton Research 16(5),
527-541.
- Spaulding S. A., McKnight D. M., Stoermer E. F., and Doran P. T. (1997) Diatoms in sediments of
perennially ice-covered Lake Hoare, and implications for interpreting lake history in the McMurdo Dry
Valleys of Antarctica. Journal of Paleolimnology 17, 403-420.
- Spigel R. H. and Priscu J. C. (1996) Evolution of temperature and salt structure of Lake Bonney, a
chemically stratified Antarctic lake. Hydrobiologia 321, 177-190.
- Toxey J. K., Meese D. A., Welch K. A., and Lyons W. B. (1997) The measurement of reactive silica
in saline-hypersaline lakes: examples of the problem. International Journal of Salt Lake Research
6, 1-7.
- Van Der Knaap E., Rodriguez R. J., and Freckman D. W. (1993) Differentiation of bacterial-feeding
nematodes in soil ecological studies by means of arbitrarily-primed PCR. Soil Biology and
Biochemistry 25(9), 1141-1151.
- Wang L. and J.C.Priscu. (1994) Influence of phytoplankton on the response of bacterioplankton growth
to nutrient enrichment. Freshwater Biology 31, 183-190.
- Wang L. and Priscu J. C. (1994) Stimulation of aquatic bacterial activity by cyanobacteria.
Hydrobiologia 277, 145-148.
- Ward B. B. and Priscu J. C. (1997) Detection and characterization of denitrifying bacteria from a
permanently ice-covered antarctic lake. Hydrobiologia 347, 57-68.
- Welch K. A., Lyons W. B., Graham E., Neumann K., Thomas J. M., and Mikesell D. (1996) Determination
of major element chemistry in terrestrial waters from Antarctica by ion chromatography. Journal of
Chromatography 739, 257-263.
- Wharton Jr. R. A., Crosby J., McKay C. P., and Rice J. (1995) Paleolakes on Mars. Journal of
Paleolimnology 13, 267-283.
- Wharton Jr. R. A., Lyons W. B., and Des Marais D. J. (1993) Stable isotopic biogeochemistry of
carbon and nitrogen in a perennially ice-covered Antarctic lake. Chemical Geology 107, 159-172.
- Yeates G. W., Bongers T., DeGoede R. G. M., Freckman D. W., and Georgieva S. S. (1993) Feeding
habits in soil nematode families and genera - an outline for soil ecologists. Journal of Nematology
25(3), 315-331.
In Press
- Lewis K. J., Fountain A. G., and Dana G. L. (in press) Surface energy balance and meltwater production for a
Dry Valley glacier, Taylor Valley, Antarctica. Annals of Glaciology.
- Lewis K. J., Fountain A. G., and Langevin P. (in press) The role of terminus cliff melt in stream flow,
Taylor Valley, McMurdo Dry Valleys. Global and Planetary Change.
- Lyons W. B., Welch K. A., and Sharma P. (in press) Chlorine-36 in the waters of the McMurdo Dry Valley Lakes,
Southern Victoria Land, Antarctica: Revisited. Geochimica et Cosmochimica Acta.
- McKnight D. M., Boyer E. W., Doran P. T., Westerhoff P. K., Kulbe T., and Andersen D. T. (in press)
Spectrofluorometric characterization of aquatic fulvic acid for determination of precursor organic material and
general structural properties. Limnology and Oceanography.
- McKnight D. M., Howes B. L., Spaulding S. A., Taylor C. D., and Goehringer D. D. (in press) Phytoplankton
dynamics in a stably stratified Antarctic lake during winter darkness. Journal of Phycology.
- Neumann K., Lyons W. B., and DesMarais D. J. (in press) Inorganic carbon isotope distribution and budget in
the Lake Hoare and Lake Fryxell basins, Taylor Valley, Antarctica. Annals of Glaciology.
- Powers L. E., Ho M., Freckman D. W., and Virginia R. A. (in press) Distribution, community structure, and
microhabitats of soil biota along an elevational gradient in Taylor Valley, Antarctica. Arctic and Alpine
Research.
- Runkel R. L., McKnight D. M., and Andrews E. D. (in press) A variable flow model of hyporheic interactions
in Huey Creek, McMurdo Dry Valleys, Antarctica. J. North Amer. Benthol. Soc.
- Tyler S. W., Cook P. G., Butt A. Z., Thomas J. M., P.T.Doran, and Lyons W. B. (in press) Evidence of deep
circulation in two perennially ice-covered Antarctic lakes. Limnology and Oceanography.
- Vandal G. M., Mason R. P., McKnight D., and Fitzgerald W. (in press) Mercury speciation and distribution in a
polar desert lake (Lake Hoare, Antarctica) and two glacial meltwater streams. Water, Air and Soil Pollution.
- Vincent W. F., Rae R., Laurion I., Howard-Williams C., and Priscu J. C. (in press) Transparency of Antarctic
lakes to solar ultraviolet radiation. Limnology and Oceanography.
In Review
- Courtright E. M., Freckman D. W., Virginia R. A., Frisse L. M., Vida J. T., and Thomas W. K. (in review)
Nuclear and mitochondrial DNA sequence diversity in the Antarctic nematode Scottnema lindsayae. Molecular
Ecology.
- Doran P. T., Berger G., Wharton Jr. R. A., Lyons W. B., Davisson L., Southon J., and Dibb J. (in review)
Dating Quaternary lacustrine sediments on the McMurdo Dry Valleys, Antarctica. Palaeography, Palaeoclimatology,
Palaeoecology.
- Fountain A. G., Lewis K. J., Dana G. L., and Doran P. T. (in review) Spatial variation in glacier mass
balances, Taylor Valley, Antarctica. Global and Planetary Change.
- Kepner Jr. R., Wharton Jr. R., Doran P. T., Kortyna A., Andersen D., and Roberts E. (in review) Effects of
research diving on a stratified antarctic lake. Limnology and Oceanography.
- Kepner Jr. R. L., Wharton Jr. R. A., and Suttle C. A. (in review) Viruses in antarctic lakes. Limnology
and Oceanography.
- Lewis K. J., Fountain A. G., and Dana G. L. (in review) How important is terminus cliff melt? A study of
the Canada Glacier terminus, Taylor Valley, Antarctica. Global and Planetary Change.
- Lyons W. B., Benson L. V., Bullen T. D., Thomas J. M., and Welch K. A. (in review-a) Strontium isotopic
signatures of the Taylor Valley Lakes, Southern Victoria Land, Antarctica: Palaeoclimatic significance. Aquatic
Geochemistry.
- Lyons W. B., Fountain A. G., Doran P. T., Priscu J. C., and Neumann K. (in review-b) The importance of
landscape position and legacy: the evolution of the Taylor Lake Valley District, Antarctica. Freshwater
Biology.
- Lyons W. B., Tyler S. W., Wharton Jr. R. A., and McKnight D. M. (in review-c) Paleoclimate history of the
McMurdo Dry Valleys, Antarctica as derived from lacustrine data. Antarctic Science.
- Lyons W. B., Tyler S. W., Wharton Jr. R. A., McKnight D. M., and Vaughn B. H. (in review-a) A Late
Holocene Desiccation of Lake Hoare and Lake Fryxell, Dry Valleys Antarctica, Derived from Lacustrine Isotope Data.
Antarctic Science.
- McKnight D. M., Runkel R., Duff J., Tate C. M., and Moorhead D. L. (in review) Nutrient uptake by algal mats
in Antarctic glacial meltwater streams. in USGS review for submission to J. North Americ. Benthol. Soc.
- McKnight D. M., Tate C. M., and Nyogi D. K. (in review) Cryogenic preservation of cyanobacterial mats in an
Antarctic stream. In USGS review for submission to Science.
- Neumann K., Lyons W. B., and Priscu J. C. (in review) CO2 Concentrations in Perenially Ice-Covered Antarctic
Lakes of Taylor Valley, Antarctica. Limnology and Oceanography.
- Takacs C. D. and Priscu J. C. (in review) Bacterioplankton dynamics in the McMurdo Dry Valley lakes:
Production and biomass loss over four seasons. Microbial Ecology.
Book Chapters
- Adams E. E., Priscu J. C., Fritsen C. H., Smith S. R., and Brackman S. L. (1998) Permanent Ice Covers of
the McMurdo Dry Valleys Lakes, Antarctica: Bubble Formation and Metamorphism. In Ecosystem Dynamics in a Polar
Desert: The McMurdo Dry Valleys, Antarctica (ed. J. C. Priscu), pp. 281-295. American Geophysical Union.
- Blair J. M., Bohlen P. J., and Freckman D. W. (1996) Soil invertebrates as indicators of soil quality. In
Methods for Assessing Soil Quality (ed. J. W. Doran and A. J. Jones), pp. 273-291. Soil Science Society of
America.
- Conovitz P. A., McKnight D. M., MacDonald L. H., Fountain A. G., and House H. R. (1998) Hydrologic
Processes Influencing Streamflow Variation in Fryxell Basin, Antarctica. In Ecosystem Dynamics in a Polar Desert:
The McMurdo Dry Valleys, Antarctica (ed. J. C. Priscu), pp. 93-108. American Geophysical Union.
- Dana G. L., Wetzel M. A., and Wharton Jr. R. A. (1996) Satellite-derived surface temperatures in the
McMurdo Dry Valleys, Antarctica. In International Radiation Symposium '96: Current Problems in Atmospheric
Radiation (ed. Smith and Stamnes), pp. 530-533. A. Deepak Publishing.
- Dana G. L., Wharton Jr. R. A., and Dubayah R. (1998) Solar Radiation in the McMurdo Dry Valleys, Antarctica.
In Ecosystem Dynamics in a Polar Desert: The McMurdo Dry Valleys, Antarctica (ed. J. C. Priscu), pp.
39-64. American Geophysical Union.
- Fountain A. G., Dana G. L., Lewis K. J., Vaughn B. H., and McKnight D. M. (1998) Glaciers of the McMurdo
Dry Valleys, Southern Victoria Land, Antarctica. In Ecosystem Dynamics in a Polar Desert: The McMurdo Dry Valleys,
Antarctica (ed. J. C. Priscu), pp. 65-76. American Geophysical Union.
- Freckman D. W. and Virginia R. A. (1998) Soil Biodiversity and Community Structure in the McMurdo Dry
Valleys, Antarctica. In Ecosystem Dynamics in a Polar Desert: The McMurdo Dry Valleys, Antarctica (ed. J.
C. Priscu), pp. 323-335. American Geophysical Union.
- Fritsen C. H., Adams E. E., McKay C. M., and Priscu J. C. (1998) Permanent Ice Covers of the McMurdo Dry
Valley Lakes: Liquid Water Contents. In Ecosystem Dynamics in a Polar Desert: The McMurdo Dry Valleys,
Antarctica (ed. J. C. Priscu), pp. 269-280. American Geophysical Union.
- Graham E. Y., Ramsey L. A., Lyons W. B., and Welch K. A. (1997) Determination of Rare Earth Elements in
Antarctic Lakes and Streams of Varying Ionic Strengths. In Plasma Source Mass Spectrometry: Developments and
Applications (ed. G. Holland and S. D. Tanner), pp. 253-262. Royal Society of Chemistry.
- Howard-Williams C., Schwarz A.-M., Hawes I., and Priscu J. C. (1998) Optical Properties of the McMurdo Dry
Valley Lakes, Antarctica. In Ecosystem Dynamics in a Polar Desert: The McMurdo Dry Valleys, Antarctica (ed.
J. C. Priscu), pp. 189-204. American Geophysical Union.
- Kepner Jr. R. L., Galchenko V., and Wharton Jr. R. A. (1997) The abundance of planktonic virus-like
particles in antarctic lakes. In Ecosystem Processes in Antarctic Ice-free Landscapes (ed. W. B. Lyons,
C. Howard-Williams, and I. Hawes), pp. 241-250. Balkema Press.
- Lizotte M. P. and Priscu J. C. (1998) Distribution, Succession and Fate of Phytoplankton in the McMurdo Dry
Valley Lakes of Antarctica, based on Pigment Analysis. In Ecosystem Dynamics in a Polar Desert: The McMurdo Dry
Valleys, Antarctica (ed. J. C. Priscu), pp. 229-240. American Geophysical Union.
- Lyons W. B., Mayewski P. A., Bartek L. R., and Doran P. T. (1997) Climate history of the McMurdo Dry
Valleys since the last glacial maximum: A Synthesis. In Ecosystem Processes in Antarctic Ice-free Landscapes
(ed. W. B. Lyons, C. Howard-Williams, and I. Hawes), pp. 15-22. Balkema Press.
- Lyons W. B., McKnight D. M., and 26 co-authors. (in press) Global Environmental Change, Catchments and
Isotopes. In Isotope Tracers in Catchment Hydrology (ed. J. McDonnell and C. Kendall).
- Lyons W. B., Welch K. A., Neumann K., Toxey J. K., McArthur R., Williams C., McKnight D. M., and Moorhead D.
(1998) Geochemical Linkages Among Glaciers, Streams and Lakes Within the Taylor Valley, Antarctica. In Ecosystem
Dynamics in a Polar Desert: The McMurdo Dry Valleys, Antarctica (ed. J. C. Priscu), pp. 77-92. American
Geophysical Union.
- Lyons W. B., Welch K. A., Nezat C. A., McKnight D. M., Crick K., Toxey J. K., and Mastrine J. A. (1997)
Chemical weathering rates and reactions in the Lake Fryxell Basin, Taylor Valley: comparison to temperate river
basins. In Ecosystem Processes in Antarctic Ice-free Landscapes (ed. W. B. Lyons, C. Howard-Williams, and I.
Hawes), pp. 147-154. Balkema Press.
- McKnight D. M., Aiken G. R., Andrews E. D., Bowles E. C., and Harnish R. A. (1993) Dissolved organic
material in dry valley lakes: A comparison of Lake Fryxell, Lake Hoare, and Lake Vanda. In Physical and
Biogeochemical Processes in Antarctic Lakes (ed. W. J. Green and E. I. Friedmann), pp. 119-133. American
Geophysical Union.
- McKnight D. M., Alger A., Tate C. M., Shupe G., and Spaulding S. (1998) Longitudinal Patterns in Algal
Abundance and Species Distributon in Meltwater Streams in Taylor Valley, Southern Victoria Land, Antarctica. In
Ecosystem Dynamics in a Polar Desert: The McMurdo Dry Valleys, Antarctica (ed. J. C. Priscu), pp. 109-128.
American Geophysical Union.
- Moorhead D. and Priscu J. C. (1998) The McMurdo Dry Valley Ecosystem: Organization, Controls and Linkages.
In Ecosystem Dynamics in a Polar Desert: The McMurdo Dry Valleys, Antarctica (ed. J. C. Priscu), pp. 351-363.
American Geophysical Union.
- Moorhead D. L., Davis W. S., and Wharton Jr. R. A. (1997) Carbon dynamics of aquatic microbial mats in the
antarctic dry valleys: A modelling synthesis. In Ecosystem Processes in Antarctic Ice-Free Landscapes (ed.
W. B. Lyons, C. Howard-Williams, and I. Hawes), pp. 181-196. Balkema Press.
- Moorhead D. L., McKnight D. M., and Tate C. M. (1998) Modeling Nitrogen Transformations in Dry Valley
Streams, Antarctica. In Ecosystem Dynamics in a Polar Desert: The McMurdo Dry Valleys, Antarctica (ed. J.
C. Priscu), pp. 141-151. American Geophysical Union.
- Neale P. J. and Priscu J. C. (1998) Fluorescence Quenching in Phytoplankton of the McMurdo Dry Valley Lakes
(Antarctica): Implications for the Structure and Function of the Photosynthetic Apparatus. In Ecosystem Dynamics
in a Polar Desert: The McMurdo Dry Valleys, Antarctica (ed. J. C. Priscu), pp. 241-254. American Geophysical
Union.
- Niles R. K. and Freckman D. W. (in press) From the ground up: nematode ecology in bioassessment and
ecosystem health. In Plant-Nematode Interactions, Agronomy Monograph (ed. K. R. Barker, G. A. Pederson, and G.
L. Windham). American Society of Agronomy, Crop Science Society of America and Soil Science Society of America.
- Niyogi D. K., Tate C. M., McKnight D. M., Duff J. H., and Alger A. S. (1997) Species composition and
primary production of algal communities in Dry Valley streams in Antarctica: examination of the functional role of
biodiversity. In Ecosystem Processes in Antarctic Ice-free Landscapes (ed. W. B. Lyons, C.
Howard-Williams, and I. Hawes), pp. 171-180. Balkema Press.
- Priscu J. C. and Sullivan C. W. (in press) Nitrogen Metabolism in Antarctic Fast-Ice Microalgal Assemblages.
In The Biology of Antarctic Sea Ice (ed. M. Lizotte and K. Arrigo). American Geophysical Union.
- Spaulding S. A. and McKnight D. M. (in press) Diatoms as indicators of environmental change in Antarctic
freshwaters. In The Diatoms: Applications to the Environmental and Earth Sciences (ed. E. F. Stoermer and J.
P. Smol). Cambridge University Press.
- Spigel R. H. and Priscu J. C. (1998) Physical limnology of the McMurdo Dry Valleys Lakes. In Ecosystem
Dynamics in a Polar Desert: The McMurdo Dry Valleys, Antarctica (ed. J. C. Priscu), pp. 153-188. American
Geophysical Union.
- Voytek M. A., Ward B. B., and Priscu J. C. (1998) The Abundance of Ammonium-Oxidizing Bacteria in Lake
Bonney, Antarctica Determined by Immunofluorescence, PCR, and In Situ Hybridization. In Ecosystem Dynamics in a
Polar Desert: The McMurdo Dry Valleys, Antarctica (ed. J. C. Priscu), pp. 217-228. American Geophysical
Union.
- Wall D. H. and Virginia R. A. (in press) The world beneath our feet: soil biodiversity and ecosystem
functioning. In Nature and Human Society (ed. P. Raven and T. A. Williams). National Academy of Sciences
Press.
- Wharton Jr. R. A. (1994) Stromatolitic mats in Antarctic lakes. In Stromatolitic mats in Antarctic
lakes (ed. J. Bertrand-Sarfati and C. Monty). Kluwer.
- Wharton Jr. R. A., McKay C. P., Clow G. D., and Anderson D. T. (1993) Perennial ice covers and their
influence on antarctic lake ecosystems. In Physical and Biogeochemical Processes in Antarctic Lakes (ed. W.
J. Green and E. I. Friedmann), pp. 53-70. American Geophysical Union.
Antarctic Journal of the United States
- Adams E. A. and Priscu J. C. (in press) Some metamorphic processes in the lake ice in the McMurdo Dry
Valleys. Antarctic Journal of the United States.
- Alger A. S., Spaulding S. A., Shupe G. H., and McKnight D. M. (in press) Species composition and spatial
distribution in Green Creek, Taylor Valley, Antarctica. Antarctic Journal of the United States.
- Booth N., Kasmer D., and McKnight D. (in press) Characteristics of streams in the Garwood Valley, McMurdo Dry
Valleys. Antarctic Journal of the United States.
- Burkins M. B., Chamberlain C. P., Virginia R. A., and Freckman. D. W. (in press) The natural abundance of
carbon and nitrogen isotopes in potential sources of organic matter to soils of Taylor Valley, Antarctica.
Antarctic Journal of the United States.
- Butt A. and Hastings J. T. (1994) McMurdo LTER: Developing a GIS Data Base, 1994. Antarctic Journal of
the United States 29(5), 244-245.
- Courtright E. M., Freckman D. W., Powers L. E., Ho M., and Virginia R. A. (in press) McMurdo LTER: Genetic
diversity of soil nematodes in the McMurdo Dry Valleys of Antarctica. Antarctic Journal of the United
States.
- Dana G. L., Fountain A. G., and Wharton Jr. R. A. (in press) McMurdo Dry Valleys LTER: Solar radiation on
glaciers in Taylor Valley, Antarctica. Antarctic Journal of the United States.
- Dana G. L., Tate C. M., and Dewey S. L. (1994) McMurdo LTER: Using narrow band spectroradiometry to assess
algal and moss communities in a dry valley stream. Antarctic Journal of the United States 29(5), 232-234.
- Doran P. T., Dana G. L., Hastings J. T., and Wharton Jr. R. A. (1995) The McMurdo LTER Automatic Weather
Network (LAWN). Antarctic Journal of the United States 30(5), 276-280.
- Doran P. T., Wharton Jr. R. A., and Schmok J. (in press) Geophysical determination of bathymetry and
morphometry of Taylor Valley lakes. Antarctic Journal of the United States.
- Doran P. T., Wharton Jr. R. A., Spaulding S. A., and Foster J. S. (1994) McMurdo LTER: Paleolimnology of
Taylor Valley. Antarctic Journal of the United States 29(5), 234-237.
- Downes M. T. and Priscu J. C. (in press) Profiles of electrode potential and dissolved oxygen in lakes of
the McMurdo dry valleys. Antarctic Journal of the United States.
- Edwards R. L. and Priscu J. C. (in press) Relationships between vertical nutrient flux and phytoplankton
biomass and productivity in lakes of the Taylor Valley, Antarctica. Antarctic Journal of the United States.
- Fountain A. G., Lewis K. J., and Dana G. L. (in press) Spatial variation of glacier mass balance in Taylor
Valley, Antarctica. Antarctic Journal of the United States.
- Fountain A. G., Vaughn B. H., and Dana G. L. (1994) Glacier mass balances of Taylor Valley, Antarctica.
Antarctic Journal of the United States 29(5), 226-228.
- Freckman D. W. and Virginia R. A. (1993) The ecology of nematodes in Antarctic Dry Valley soils.
Antarctic Journal of the United States 28, 10-11.
- Hastings J. T. (in press) Progress with the Taylor Valley GIS 1995. Antarctic Journal of the United
States.
- Ho M., Virginia R. A., Powers L. E., and Freckman D. W. (1995) Soil chemistry along a glacial chronosequence
on Andrews Ridge, Taylor Valley, Antarctica. Antarctic Journal of the United States 30, 310-311.
- Kasmer D., Booth N., and McKnight D. (in press) A comparison of two separate visits to the Alph River system
with the Onyx River system. Antarctic Journal of the United States.
- Kepner Jr. R. L. and Wharton Jr. R. A. (in press) McMurdo LTER: Characterization of protozoan communities
in Lakes Hoare and Fryxell utilizing artificial substrates. Antarctic Journal of the United States.
- Lewis K., Fountain A., and Langevin P. (in press) The role of terminus cliff melt in the hydrological cycle,
Taylor Valley, Antartica. Antarctic Journal of the United States.
- Lewis K. L., Dana G., Fountain A., and Tyler S. (1995) The surface energy balance of the Canada Glacier,
Taylor Valley. Antarctic Journal of the United States 30(5), 280-282.
- Lyons W. B., Tyler S. W., Welch K. A., and Sharma P. (in press) Density-driven mixing in Lake Hoare?
Antarctic Journal of the United States.
- McKnight D., House H., and Von Guerard P. (1994) McMurdo LTER: Streamflow measurements in Taylor Valley.
Antarctic Journal of the United States 29, 230-231.
- McKnight D. M. and Tate C. M. (in press) Algal mat distribution in glacial meltwater streams in Taylor
Valley, Southern Victoria Land, Antarctica. Antarctic Journal of the United States.
- Moorhead D. L. and McKnight D. (in press) Stream discharge as a function of ambient temperature and incoming
shortwave radiation in Taylor Valley, Antarctica. Antarctic Journal of the United States.
- Moorhead D. L. and Wharton Jr. R. A. (1994) Primary production model of benthic microbial mats in Lake
Hoare, Antarctica. Antarctic Journal of the United States 29(5), 241-243.
- Powers L. E., Freckman D. W., Ho M., and Virginia R. A. (1994) McMurdo LTER: Soil and nematode distribution
along an elevational gradient in Taylor Valley, Antarctica. Antarctic Journal of the United States 29(5),
228-229.
- Powers L. E., Freckman D. W., Ho M., and Virginia R. A. (1995) McMurdo LTER: Soil properties associated
with nematode distribution along an elevational transect in Taylor Valley, Antarctica. Antarctic Journal of the
United States 30, 282-283.
- Powers L. E., Freckman D. W., and Virginia R. A. (1994) Depth distribution of soil nematodes in Taylor
Valley, Antarctica. Antarctic Journal of the United States 29(5), 175-176.
- Priscu J. C. (1994) Phytoplankton nutrient deficiency in lakes of the McMurdo Dry Valleys, Antarctica.
Antarctic Journal of the United States 29(5), 239-240.
- Priscu J. C. (1995) Phototactic response of phytoplankton forming discrete layers within the water column of
Lake Bonney, Antarctica. Antarctic Journal of the United States 30, 301-302.
- Takacs C. D. and Priscu J. C. (1995) Responses of bacterial growth to inorganic and organic nutrient
enrichment in the lakes of the Dry Valleys, Antarctica. Antarctic Journal of the United States 30, 303-304.
- Takacs C. D. and Priscu J. C. (in press) The role of phytoplankton extracellular release in bacterioplankton
growth of Taylor Valley lakes, Antarctica. Antarctic Journal of the United States.
- Welch K. A. and Lyons W. B. (in press) Comparative limnology of the Taylor Valley Lakes: The major solutes.
Antarctic Journal of the United States.
- Welch K. A., Lyons W. B., Priscu J. C., Edwards R., McKnight D. M., House H., and Wharton Jr. R. A.
(1994) McMurdo LTER: Inorganic geochemical studies with special reference to CaCO3 dynamics. Antarctic Journal
of the United States 29(5), 237-238.
- Wharton Jr. R. A. (1994) McMurdo Dry Valleys Long-Term Ecological Research (LTER): An overview of 1993-94
research activities. Antarctic Journal of the United States 29(5), 224-225.
Theses/Dissertations
- Courtright E. M. (1995) Soil nematode distribution and genetic diversity in the Dry Valleys of Antarctica.
Master's Thesis, Colorado State University, Fort Collins, CO.
- Davis W. S. (1998) Spatial-temporal patterns of primary production in phytoplankton communities of Taylor
Valley lakes, Antartica: A modelling synthesis. Master's Thesis, Texas Tech University.
- Doran P. T. (1996) Paleolimnology of Perennially Ice-Covered Antarctic Oasis Lakes. Ph.D. Dissertation,
University of Nevada, Reno.
- Lewis K. J. (1996) Surface energy balance and meltwater production for a dry valley glacier, Taylor Valley,
Antarctica. Master's Thesis, University of Colorado.
- Neumann K. (1998) Inorganic Carbon Cycling in Perennially Ice-Covered Lakes in Taylor Valley, Antartica. Ph.D.
Dissertation, University of Alabama, Tuscaloosa, AL.
- Nezat C. A. (1998) Chemical Weathering in Taylor Valley, Antarctica: Quantity and Quality. Master's Thesis,
University of Alabama, Tuscaloosa, AL.
- Spaulding S. A. (1995) Algal investigations at varying temporal scales in an extreme environment: McMurdo Dry
Valley lakes, Antarctica. Ph.D. Dissertation, Colorado State University, Fort Collins, CO.
Other Publications
- Alger A. S., McKnight D. M., Spaulding S. A., Tate C. M., Shupe G. H., Welch K. A., Edwards R., Andrews E.
D., and House H. R. (1997) Ecological Processes in a Cold Desert Ecosystem: The Abundance and Species Distribtuon
ofAlgal Mats in Glacial Meltwater Streams in Taylor Valley, Antarctica. Occasional Paper No. 51, 102 pp.
Institute of Arctic and Alpine Research, University of Colorado, Boulder, CO.
- Bargagli R., Wynn-Williams D., Bersan F., Cavacini P., Ertz S., Frati F., Freckman D. W., Lewis-Smith R.,
Russell N., and Smith A. (1997) Field Report BIOTEX I: First BIOTAS Expedition (Edmonson Point-Bais Terra Nova,
Dec 10, 1995-Feb 6, 1996). In Newsletter for the Italian Biological Research in Antarctica (ed. M.
Tamburrini and R. D'Avino). Universita degli Studi di Camerino, Gennaio, Italy.
- Brown M. W. (1995) Antarctica may offer clues on whether life existed on Mars. In New York Times, pp.
Sci B9.
- Doran P. T. (1996) A weather network in McMurdo Dry Valleys, Antarctica. Campbell Scientific, Inc.
- Freckman D. W. (1994) Life in the soil. Soil biodiversity: its importance to ecosystem processes. Report of
a Workshop held at The Natural History Museum, London, England. pp. 26.
- Freckman D. W. (in press) Soil Biodiversity: Life in Soil. In The Living Planet in Crisis (ed. J.
Cracraft and F. Griffo). Columbia University Press.
- McKnight D. M. (ed.) (1997) McMurdo Dry Valley LTER Science Workshop. Institute of Arctic and Apline
Research, Boulder, Colorado, pp. 50.
- Priscu J. C. (1994) Book Review: Physical and biogeochemical processes in Antarctic lakes. (1993) Green and
Friedmann (eds.). Limnology and Oceanography 39(6), 1499-1500.
- Von Guerard P., McKnight D. M., Harnish R. A., Gartner J. W., and Andrews E. D. (1994) Streamflow,
water-temperature, and specific conductance data for selected streams draining into Lake Fryxell, lower Taylor
Valley, Victoria Land, Antarctica, 1990-92. United States Geological Survey, Denver.
Abstracts
- Bamforth S. S., Freckman D. W., and Virginia R. (1995) Early soil biogenesis. Soil Ecology Society of
America Annual Meeting, Fort Collins, CO.
- Bamforth S. S., Freckman D. W., and Virginia R. A. (1996) Amoebae biodiversity in the Antarctic Dry Valley
Soils. Seventh International Conference on Small Freeliving Amoebae, Adelaide.
- Brown M. J., Virginia R. A., and Chamberlain C. P. (1996) Sources and distribution of abundance of organic
matter in the Dry Valley soils of Southern Victoria Land, Antarctica. Ecological Society of America Annual
Meeting, 77. Providence, RI.
- Burkins M. B., Virginia R. A., Chamberlain C. P., and Freckman D. W. (1997) Carbon cycling in the soils of
the McMurdo Dry Valley region, Antarctica. Ecological Society of America Annual Meeting, Albuquerque, NM.
- Dana G. L. and Wharton Jr R. A. (1996) Radiation balance in the McMurdo Dry Valleys, Antarctica.
American Geophysical Union Fall Meeting, San Francisco, CA.
- Dana G. L., Wharton Jr. R. A., and Tyler S. W. (1997) Surface energy balance of perennially ice-covered
lakes in the McMurdo Dry Valleys, Antarctica. Aquatic Sciences meeting of the American Society of Limnology and
Oceanography, Santa Fe, NM.
- Downes M. T. and Priscu J. C. (1995) The biogeochemistry of nitrogen in the water column of Lake Bonney, an
Antarctic dry valley lake. Pacific Chemistry Conference (PACIFICHEM), Honolulu, HI.
- Downes M. T. and Priscu J. C. (1996) The biogeochemistry of nitrogen in the water column of Lake Boney, an
Antarctic dry valley lake. Polar Desert Ecosystems International Workshop, Christchurch, New Zealand.
- Edwards R. L. and Priscu J. C. (1995) Relationship between vertical nutrient flux and phytoplankton biomass
and productivity in lakes of the McMurdo Dry Valleys. American Society of Limnology and Oceanography Annual
International Meeting, Reno, NV.
- Edwards R. L. and Priscu J. C. (1997) Seasonal development of photosynthesis and biomass in lakes of the
McMurdo Dry Valleys, Antarctica. American Society of Limnology and Oceanography Annual International
Meeting, Sante Fe, NM.
- Freckman D. W. and Virginia R. A. (1997) Soil nematode communities and disturbance: A perspective from
different deserts. Ecological Society of America Annual Meeting, Albuquerque, NM.
- Freckman D. W., Virginia R. A., and Powers L. E. (1994) Nematode biodiversity and survival in Antarctic Dry
Valley soils. Scientific Committee for Antarctic Research (SCAR) Sixth Biology Symposium, Venice, Italy.
- Ho M., Virginia R. A., and Freckman D. W. (1996) Soil spatial variation along a toposequence in Taylor
Valley, Antarctica. Ecological Society of America Annual Meeting, Providence, RI.
- Kepner Jr. R., Galchenko V., and Wharton Jr. R. A. (1997) The abundance of planktonic viruses in antarctic
lakes. First International Workshop on Polar Desert Ecosystems, The Netherlands.
- Kepner Jr. R. L. and Wharton Jr. R. A. (1997) Abundance and production of viruses in antarctic lakes.
American Society of Limnology and Oceanography Annual International Meeting, Santa Fe, NM.
- Lewis K. J., Fountain A. G., and Dana G. L. (1997) How important is terminus cliff melt?: A study of the
Canada Glacier terminus, Taylor Valley, Antarctica. IAMAS/IAPSO Conference, Melbourne, Australia.
- Lewis K. J., Fountain A. G., and Dana. G. L. (1997) Surface energy balance and meltwater production for a
dry valley glacier, Taylor Valley, Antarctica. Antarctic CRC Conference, Antarctica and Global Change:
Interactions and Impacts, Hobart, Tasmania, Australia.
- Lizotte M. P. and Priscu J. C. (1995) Phytoplankton population dynamics in Antartic lakes, as measured by
pigment analysis. American Society of Limnology and Oceanography, Reno, NV.
- Lyons W. B., Welch K. A., Graham E. Y., Priscu J. C., Benson L. V., Bullen T. D., and Green W. J. (1997)
The Geochemistry of and Chemical Evolution of Lakes Bonney and Fryxell, Antarctica: The Angino Legacy.
Geological Society of America Annual Meeting, Salt Lake City, UT.
- Neumann K., Lyons W. B., DesMarais D. J., Priscu J. C., and Edwards R. L. (1995) 13C and pCO2 distribution
in Antarctic lakes: Ice cover and biota leave their imprint. American Geophysical Union Fall Meeting, San
Francisco, CA.
- Nezat C. A., Lyons W. B., Graham E. Y., Welch K. A., Lechler P. J., and McKnight D. M. (1997) Chemical
Weathering in Streams from Taylor Valley, Antarctica. Geological Society of America Annual Meeting, Salt
Lake City, UT.
- Powers L. E., Freckman D. W., and Virginia R. A. (1996) Effects of human disturbance on soil nematode
populations in Taylor Valley, Antarctica. Ecological Society of America Annual Meeting, Providence, RI.
- Priscu J. C. and Dore J. E. (1997) Nutrient deficiency in lakes of the McMurdo Dry Valleys, Antarctica.
American Society of Limnology and Oceanography Annual Meeting, Santa Fe, NM.
- Priscu J. C. and Neale P. J. (1995) Photoactic response of phytoplankton forming discrete layers within the
water column of Lake Bonney, Antarctica. American Society of Limnology and Oceanography, Reno, NV.
- Priscu J. C., Sharp T. R., and Lizotte M. P. (1994) Development of phytoplankton structure and
photosynthesis during the winter-spring transition in Lake Bonney, a permanently ice-capped lake in the dry valleys
of McMurdo Sound, Antarctica. Scientific Committee for Antarctic Research (SCAR) Sixth Biology Symposium,
Venice, Italy.
- Takacs C. D. and Priscu J. C. (1995) Responses of bacterial growth to inorganic and organic nutrient
enrichment in Antarctic lakes. American Society of Limnology and Oceanography Annual International Meeting,
Reno, NV.
- Takacs C. D. and Priscu J. C. (1996) Bacterial growth in Antarctic lakes: The role of phytoplankton
extracellular release. American Society of Limnology and Oceanography Annual Meeting, Milwaukee, WI.
- Treonis A. M., Freckman D. W., and Virginia R. A. (1997) Water: Limiting factor for soil nematode
communities in cold deserts? Ecological Society of America Annual Meeting, Albuquerque, NM.
- Virginia R. A. (1997) The soil ecosystems: Lessons from the extremes. Soil Ecology Society
Conference, Kansas State University, Manhattan, KS.
- Voytek M. A., Priscu J. C., and Ward B. B. (1995) Detection of ammonium oxidizing bacteria in antarctic lake
samples using the polymerase chain reaction. American Society of Limnology and Oceanography Annual Meeting,
Reno, NV.
- Ward B. B. and Priscu J. C. (1995) Detection and characterization of denitrifying bacteria in an antarctic
lake. American Society of Limnology and Oceanography Annual Meeting, Reno, NV.
- Welch K. A., Neumann K., Lyons W. B., Priscu J. C., Edwards R. L., McKnight D. M., House H., and Wharton
Jr. R. W. (1995) Inorganic geochemical study of ice-covered lakes in the Taylor Valley, Antarctica. American
Society of Limnology and Oceanography Annual Meeting, Reno, NV.
- Wharton Jr. R., Dana G., Fountain A., Freckman D., Hastings J., Lyons W. B., McKnight D., Moorhead D., Priscu
J. C., and Tate C. (1994) McMurdo Dry Valleys long term ecological research site. Scientific Committee for
Antarctic Research (SCAR) Sixth Biology Symposium, Venice, Italy.
- Wharton Jr. R. A. (1996) Antarctic dry valley lakes: Sensitive indicators of environmental change.
American Geophysical Union Annual Meeting, San Francisco, CA.
MCM-LTER Datasets Electronically Available - Field Data
| Category
|
Title
|
Short Term / Long Term Study
|
|
|
I. Bathymetry
|
|
|
|
|
|
Bathymetric Polynomials
|
ST
|
|
|
|
Bathymetry Raw Data Files
|
ST
|
|
|
|
Bathymetric Values From Contour Map Digitizing
|
ST
|
|
|
|
Depths, Areas, Volumes
|
ST
|
|
|
|
Bathymetric Hypsographic Function Values
|
ST
|
|
|
II. Geochemistry
|
|
|
|
|
|
Major Ion Concentrations for Glacier Ice, Snow and Melt Water Samples
|
LT
|
|
|
|
Major Ion Chemistry for Miscellaneous Locations Throughout Taylor Valley
|
ST
|
|
|
|
Limnological Water Column & Pore Water Cations/Anions |
ST
|
|
|
|
Limnological pHs
|
LT
|
|
|
|
Lake Nutrients
|
LT
|
|
|
|
Miscellaneous Stream Chemistry Samples Throughout Taylor Valley
|
ST
|
|
|
|
Stream Nutrients (nitrate, nitrite, ammonium, reactive phosphorus)
|
LT
|
|
|
|
Stream Chemistry / Dissolved Organic Carbon 1990-1993
|
LT
|
|
|
|
Stream Chemistry / Major Ion Concentrations
|
LT
|
|
|
|
Stream Conductivity, pH 1990-1992
|
LT
|
|
|
|
Major Ion Data for Lake Ice Samples
|
ST
|
|
|
|
Limnological Chemistry / Major Ion Concentrations
|
LT
|
|
|
III. Glaciology
|
|
|
|
|
|
Glacier Snow Densities
|
LT
|
|
|
|
Glacier Mass Changes
|
LT
|
|
|
|
Average Stake Heights, Snow Depths Found at Glacier Stakes
|
LT
|
|
|
|
Glacier Stake Heights, Snow Depths
|
LT
|
|
|
|
Canada Glacier Ice Temperatures
|
LT
|
|
|
|
Canada Glacier Eddy Correlation Data
|
ST
|
|
|
|
Glacier Mass Balance Summary
|
LT
|
|
|
IV. Hydrology
|
|
|
|
|
|
Discrete Stream Gage Measurements pre-1980 (pre-LTER)
|
LT
|
|
|
|
Mean Daily Stream Gage Measurements 1969-1997
|
LT
|
|
|
|
Discrete Stream Gage Measurements 1980-1989 (pre-LTER)
|
LT
|
|
|
|
Discrete Stream Gage Measurements 1990-1997
|
LT
|
|
|
|
Taylor Valley Water Budgets
|
LT
|
|
|
|
Field Meter Stream Measurements 1990-1997
|
LT
|
|
|
V. Limnology
|
|
|
|
|
|
Ice Thickness, Piezometric Depths for Taylor Valley Lakes
|
ST
|
|
|
|
Winter Phytoplankton 1990-1991 (pre-LTER)
|
ST
|
|
|
|
Chlorophyll-A Concentrations in Lake Hoare Benthic Mats 1996-97
|
ST
|
|
|
|
Lake Chlorophyll, Primary Productivity, Respiration
|
LT
|
|
|
|
Phytoplankton Densities
|
LT
|
|
|
|
Dissolved Oxygen, Photosynthetically Active Radiation
|
LT
|
|
|
|
Bacterial Productivity and Density
|
LT
|
|
|
|
Lake Temperature and Conductivity
|
LT
|
|
|
VI. Meteorology
|
|
|
|
|
|
Commonwealth Glacier Meteorological Station Measurements (1993-1997)
|
LT
|
|
|
|
Average Meteorological Measurements for Canada Glacier (1994-1997)
|
LT
|
|
|
|
Lake Brownsworth Meteorological Station Measurements (1994-1997)
|
LT
|
|
|
|
Lake Bonney Meteorological Station Measurements (1993-1997)
|
LT
|
|
|
|
Lake Vanda Meteorological Station Measurements (1994-1997)
|
LT
|
|
|
|
Canada Glacier Meteorological Station Measurements (1995-1996)
|
LT
|
|
|
|
Lake Hoare Meteorological Station Measurements (1993-1997)
|
LT
|
|
|
|
Lake Fryxell Meteorological Station Measurements (1993-1997)
|
LT
|
|
|
|
Lake Vida Meteorological Station Measurements (1995-1997)
|
LT
|
|
|
|
Explorer's Cove Meteorological Station Measurements (1995-1996)
|
LT
|
|
|
|
Taylor Glacier Meteorological Station Measurements (1994-1997)
|
LT
|
|
|
|
Howard Glacier Meteorological Station Measurements (1993-1997)
|
LT
|
|
|
VII. Soil Ecology
|
|
|
|
|
|
Soil Organism Responses to Long-Term Soil Manipulation
|
LT
|
|
|
|
Chlorophyll-A Responses to Long-Term Algae Amendment
|
LT
|
|
|
|
Soil Moisture Responses to Long-Term Algae Amendment
|
LT
|
|
|
|
Soil Organism Responses to Long-Term Algae Amendment
|
LT
|
|
|
|
Chlorophyll-A Responses to Long-Term Soil Manipulation
|
LT
|
|
|
|
Soil Moisture Responses to Long-Term Soil Manipulation
|
LT
|
|
|
VIII. Stream Ecology
|
|
|
|
|
|
Stream Algae Ash-Free Dry Mass (1994, 1995)
|
LT
|
|
|
|
Algal Species, Morphotype Descriptions
|
ST
|
|
|
|
Stream Algae Photosynthesis/Light Measurements (1995)
|
LT
|
|
|
|
Stream Algal and Moss Biomass (1994-1997)
|
LT
|
|
|
|
Stream Algae Primary Productivity (1995, 1996, 1997)
|
LT
|
|
|
|
Stream Invertebrate Taxa Qualitative Abundances (1994)
|
LT
|
|
|
Stream Algal Abundances (1994)
|
LT
|
|
|
IX. Study Locations
|
|
|
|
|
|
Locations, Dates, Codes for Lake Chemistry, Biology Samples
|
ST
|
|
|
|
Stream Lengths for Taylor Valley Streams
|
ST
|
|
|
|
Stream Gage Locations
|
ST
|
|
|
|
Stream Transect Locations
|
ST
|
|
|
|
Coordinate List of Stream Transect Points
|
ST
|
|
|
|
Coordinate List of Stream Gage Features
|
ST
|
|
|
|
1993-94 Geodetic Survey Results / Stream Transect Reference Marks
|
ST
|
|
|
|
GPS Data, Optical Observations Producing Coordinate Files
|
ST
|
|
|
|
Glacier Stake Locations
|
ST
|
|
|
|
1996-97 MCM-LTER GPS Coordinate Locations
|
ST
|
|
|
|
Relative Positions of Stream Transect Line Points
|
ST
|
|
|
X. Data Extraction Tools
|
|
|
|
|
|
Meteorological Data Extraction Tool
|
LT
|
|
|
|
Hydrological Data Extraction Tool
|
LT
|
|
When the database was located at DRI, the Antarctic hydrology and meteorologic
data bases were publicly available. The Antarctic hydrology website was linked
to other Antarctic websites (e.g. the ICAIR site) and USGS hydrologic websites,
in addition to the LTER site. Unfortunately, we do not have documentation for
the use of these databases by non-MCM-LTER scientists. Most other databases
have only become publicly available since the transfer of the database to
INSTAAR in the summer of 1997.